Workers not shirkers
Synopsis : Not all foragers are equal. A small proportion – the elite foragers – make the majority of foraging trips. These are the most experienced foragers. Could pathogens and pesticides that reduce worker longevity compromise nutrition of the hive?
Introduction
All bees are the same.
Right?
No, of course not.
The three castes, from The ABC of Bee Culture, 1895
For a start there are three castes of honey bee; the queen, drones and workers … but we can also sub-divide these castes.
Queens
For example, most beekeepers would agree that there are fundamental and important differences between a virgin queen and a mated queen. They behave differently, their physiology is different and so are their their senses.
Drones
Similarly, the difference between virgin and mated drones is also pretty fundamental. In fact, it’s literally a matter of life and death 😉 . However, there are also less dramatic – largely physiological – differences between sexually immature and mature drones.
Workers
And there are differences in this caste as well.
Any beekeeper who uses Pagden’s artificial swarm for their swarm control has – although perhaps unknowingly – exploited the difference between two broad groups of workers; the hive (or nurse) bees and the flying bees (or foragers). The former are bees that have yet to fly from the hive. They rear the developing brood, look after the queen and perform a range of housekeeping duties.
After about three weeks the maturing worker goes on several orientation flights and eventually becomes a forager, responsible for collecting the pollen, nectar, resin and water the colony needs.
‘Eventually’ because workers undertake additional roles e.g. guard bees, undertaker bees and scouts, as they segue from hive bees to foragers {{1}}. This change in roles during the lifetime of a worker bee is termed temporal polyethism {{2}}.
Elite foragers
In this post I’m going to focus on the last of the roles the worker fulfils, that of foraging.
There is a lot of good observational and experimental science on foraging behaviour; for example, the preference of foragers for certain pollen or nectar sources, or the features of the colony that induces foraging activity. Some of this is briefly reviewed in ’A closer look – Foraging behaviour’ by Clarence Collison in Bee Culture {{3}}.
Instead of rehashing those things I’m instead going to describe the concept of ’elite’ foragers. These are a minority of the forager population that do the majority of the foraging. They are therefore probably a very important cohort of bees for the colony.
The definition of elite foragers was first demonstrated for honey bees in studies conducted by Paul Tenczar working with Gene Robinson in 2014 {{4}}.
Gross differences in foraging activity was not a new concept. It had been observed in a wide range of eusocial insects in studies dating back to the 1970’s. Since reproductive fitness of eusocial insects – like bees, wasps and ants – is determined at the colony level, and workers are genetically related, variation in worker performance was neither expected nor had an obvious origin.
However, significant differences in worker performance are observed when suitable technology exists to detect it.
’We have the technology’ {{5}}
Tenczar used RFID tags to label individual worker bees. I’ve described this technology before. It allows the unique identification of individual bees. Foragers were detected leaving or arriving at the hive by monitoring them with two RFID ‘readers’ arranged along the narrow hive entrance tunnel.
Theoretically at least, a bee registered by the inner and then the outer reader should be leaving the hive, whereas one registered first by the outer reader, followed by the inner, would be arriving. If these two pairs of events were separated by a several minutes it should mean the bee has successfully completed a round trip.
Unfortunately, the RFID/reader technology was in its (relative) infancy {{6}} and trips were missed. They even added two RFID chips to each bee to improve detection rates. Manual observation showed that there was a 76-94% chance of a trip being detected by at least one of the four readers. They therefore used reads rather than trips as a metric for activity level. This is a bit of a fudge but it will do for the purpose of this study.
As I will show shortly, the technology has now improved and there are more accurate ways to measure foraging trips.
Orientation flights and the age of onset of foraging
Tenczar et al., labelled over 1000 day-old workers in five separate experimental colonies and monitored their activity for 5-7 weeks.
Orientation flights occur before foraging flights, and usually take place in the afternoon. To be sure they were only monitoring foraging flights, they defined the first day of foraging activity as the one when there were at least 6 ‘reads’, and with at least 25% of reads occurring before midday.
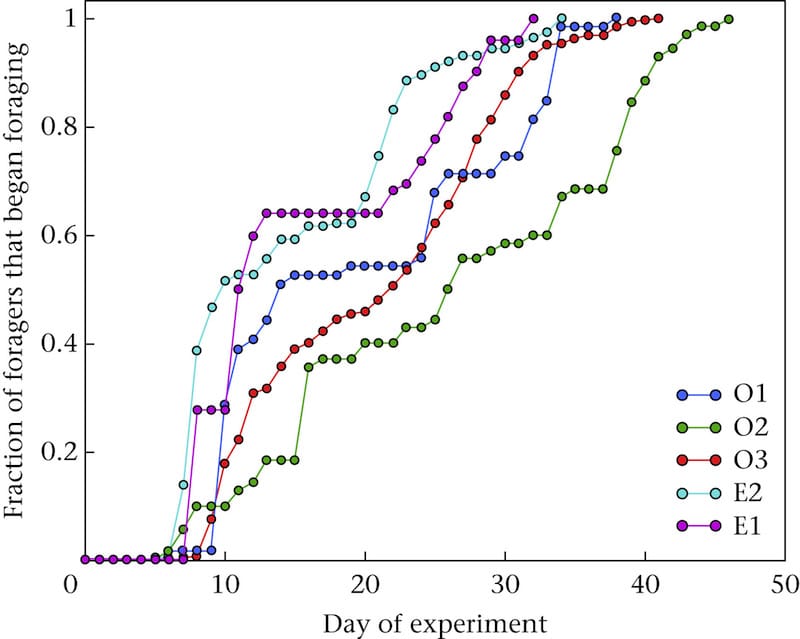
Age at onset of foraging for tagged bees
The average age of a worker at the onset of foraging was 20.4 days – a figure in agreement with the ‘three weeks in the hive’ statement I made above. However, if you average the five separate lines of hive data (above) it is also clear that a significant proportion of the bees (actually 27% of them) started foraging within the first 10 days.
This is so-called ’precocious foraging’ and had been seen previously in colonies created with a single cohort of bees. The colonies used in these studies were small (~1000 bees in each) and were started with bees of all the same age. The fact that some start foraging precociously is a demonstration of the plasticity in temporal polyethism I referred to in an earlier footnote … and which non-scientists would describe as doing ‘different jobs at different times, that might vary’.
Workers and shirkers
However, just looking at the cumulative count of workers (above) it is clear that there is considerable variation in the timing of the onset of foraging.
In addition, and perhaps more surprisingly, the level of foraging activity also varied greatly.
Some workers made rather few foraging trips, others started early and finished late, making repeated closely-spaced trips throughout the day.
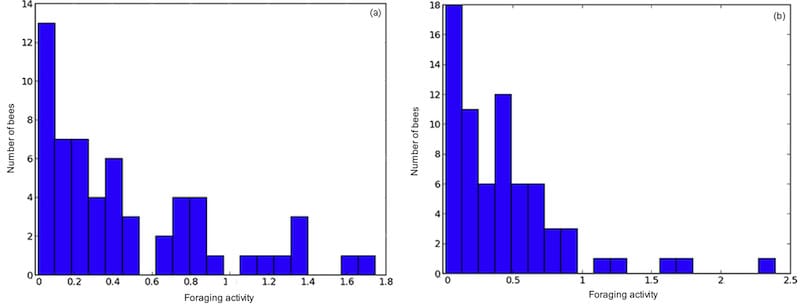
Contributions of individual bees to the total foraging activity in two colonies
These histograms show the relative foraging activity of RFID tagged bees in two representative colonies. The vertical axis records the number of bees making a particular relative foraging effort. The shape of the graph – large bars on the left and much smaller bars on the right – show that the majority of the bees (hence the large bars) do relatively little foraging, whereas a much smaller number of bees (in the shorter bars on the right) do lots.
Lorenz curves
A more informative way to represent this data is to use a Lorenz curve which displays the share of foraging activity (vertical axis) against the percentage of foraging bees (horizontal axis).
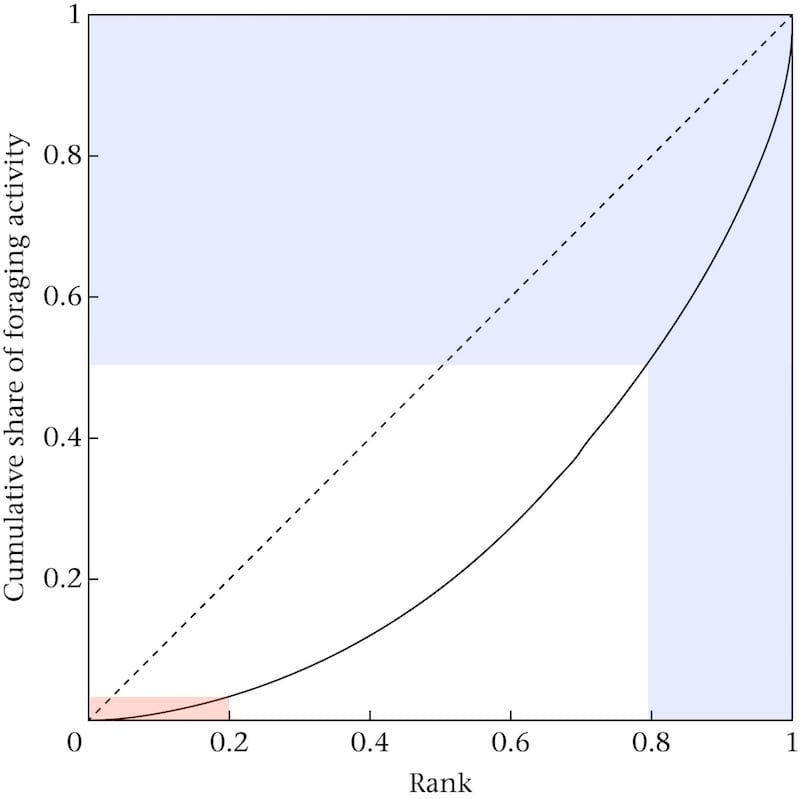
Example plot of a typical Lorenz curve of cumulative share of foraging activity for one of five study colonies
If all foragers contributed equally to foraging activity the ‘curve’ would be the diagonal dotted line i.e. 50% of the bees would ‘deliver’ 50% of the foraging activity.
In the colonies studied, the actual contribution to foraging activity is shown by the curved line.
Approximately 20% of the foragers accounted for 50% of the foraging activity (the area I’ve shaded blue). These are the elite foragers. Conversely, the ‘laziest’ 20% of foragers make less than 5% of the foraging trips (shaded red) {{7}}.
These are the shirkers … the less said about them the better 😉 .
Tenczar et al., conducted additional analysis of the pattern of foraging activity per bee per day, and the consequences of removal of elite foragers (in due course other foragers become elite foragers). However, I’ll skip these as I want to move on to a more recent study of elite foragers.
We really do ‘have the technology’
In 2019 Simon Klein and colleagues published another RFID-tagged forager study {{8}}. In the intervening years the RFID tag and reader technology had improved. They used two modified four frame nucleus hives in which bees traversed separate tunnels for entry and exit.
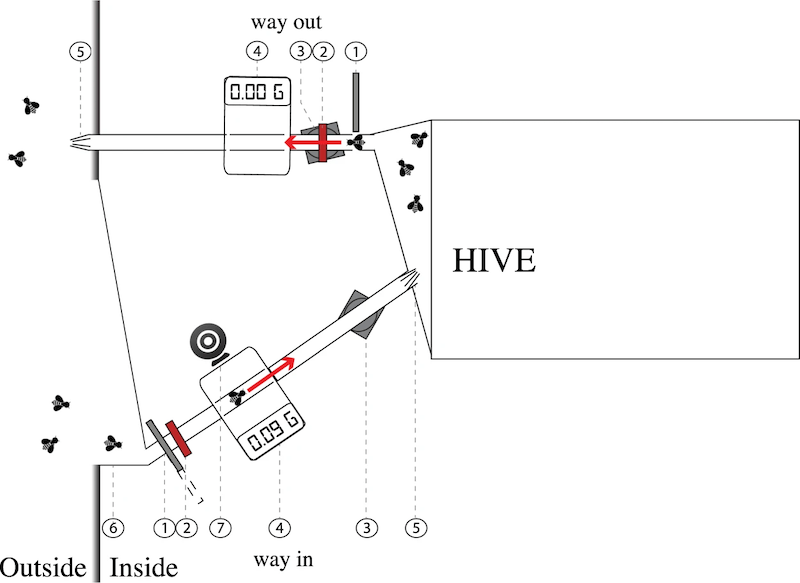
Colony entrance with sensors – bees enter and depart the hive using different tunnels
In addition to using more reliable RFID readers (#3 in the diagram above) they also weighed (#4) the bees as they entered and exited the hive and recorded video (#7) of the returning bees to determine whether they were carrying pollen.
These additional measurements meant that, in addition to the number of trips completed, the authors were also able to measure foraging performance in terms of pollen or nectar collected.
Actually, that’s a bit of an overstatement.
Pollen foragers were identifiable on video by their pollen-filled corbiculae (PDF). In contrast, foragers returning without pollen could have made unsuccessful trips, or may have collected nectar or water. They therefore classified foraging activity into ‘pollen’ or ‘non-pollen’ trips.
In total they monitored 564 foragers who made an average of 19 foraging trips in their lifetime (~10,500 trips in total). Interestingly, the average foraging lifetime was less than 5 days. As with Tenczar et al., they excluded orientation flights from the trips recorded (though in a different way).
Practice makes perfect
None of the bees monitored foraged exclusively for pollen. However, it was noted that the more trips a bee made, the greater the proportion of the trips were for pollen until a maximum was reached, after which pollen collecting declined.
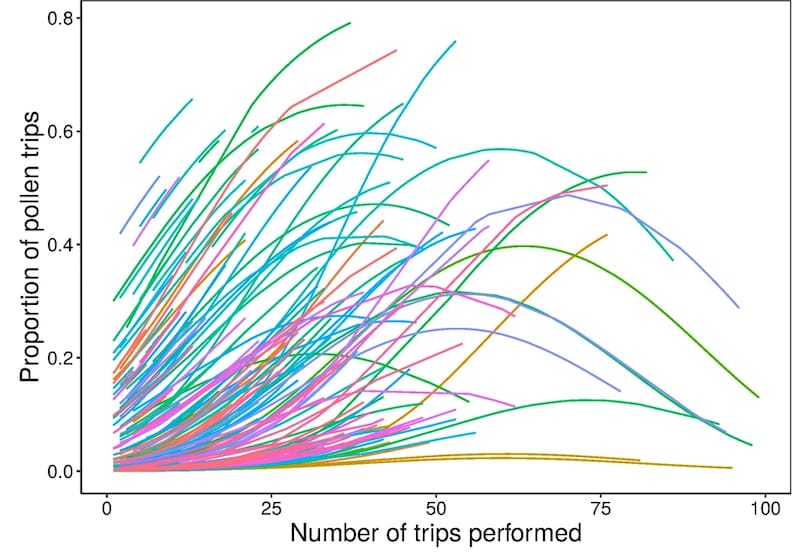
Changes of foraging performance with experience – pollen collection (each line is an individual tagged bee)
Foragers lost weight on pollen foraging trips – presumably using crop contents to ‘fuel’ the flight. Since the weight of the crop contents were unknown, it wasn’t possible to calculate the weight of the pollen collected.
Non-pollen foragers weighed about the same (or a little more) upon return as when they left. Since both the weight of the crop contents and the identity of what was being collected (water or nectar) were unknown, it was not possible to determine how much had been collected. However, as individual bees aged – and so took more non-pollen foraging trips – their gain in weight per trip increased. This suggested that (as with the likelihood of collecting pollen) increased experience resulted in more efficient foraging.
The elite foragers are the best performing foragers
Analysis of the average number of foraging trips per day demonstrated that it increased over the first ten days and then plateaued at about 10 trips per day.
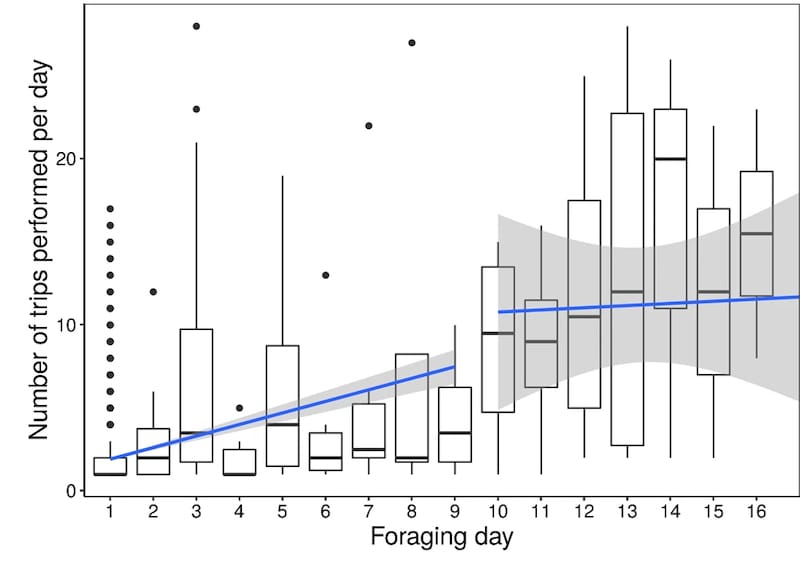
Changes of foraging performance with experience – average trips per day
It’s worth noting a couple of points here; firstly, on average foragers only made 19 trips in their lifetime and secondly, the majority of the foragers never reached maximal pollen foraging activity (in the multicoloured graph above, most lines terminate before they reach a peak and start to decline).
These points, together with the average foraging lifetime being under five days, indicate that there is a very high attrition rate amongst foragers.
Most die young … by which I mean within the first week of leaving the hive 🙁 .
But, some live long enough to become the experienced elite foragers, and these bees were the best performing foragers.
Like the previous study, an average of 19% of the foragers performed 50% of all foraging trips recorded {{9}}. These were the elite bees.
These elite bees were the most likely to collect pollen and – on non-pollen trips – were most likely to show a gain in weight, indicating a greater resource (nectar or water) load was being carried.
Conclusions and consequences
The logical conclusion is that, through experience, bees improve their foraging performance. However, most bees never realise their full potential as they perish long before they achieve the status of elite foragers.
It is known that bees exhibit both learning and memory. With regard to foraging, it’s known that navigation skills improve with experience, and that both flower discrimination and ‘handling’ also get better i.e. they are more likely to find (and re-find) remote flowers, to distinguish them from other flowers in the immediate area and to harvest the pollen or nectar from the flower.
These elite bees must make a significant contribution to resourcing the colony.
Numerically they are a minority of the foraging population, but they collect lots of the pollen, nectar, water and propolis needed by the colony.
However, without knowing the precise number and age/experience distribution of the foragers and the mass of the pollen/nectar loads collected, it is not possible to determine whether they collect the majority of these resources.
For example, do 1000 inexperienced foragers collect more (or less) than 100 elite foragers? Are the massed ranks of young naïve foragers more effective at provisioning the colony than a few dozen of ‘old timers’?
We don’t know … yet.
The experiments needed to determine this are more difficult, and a lot more intrusive. You need to know both the weight of whatever was collected, together – in the case of pollen and nectar – with its value to the colony. For example, we know that bees preferentially forage on particular high protein pollens, or on high-sucrose nectar sources … presumably (though it needs to be shown) elite bees do this better than naïve foragers.
Looking after the elderly
Tenczar et al., showed that depleting the elite bee population had little long-term effect, because younger bees made additional foraging trips in subsequent days. However, this ‘replacement’ was only measured in terms of foraging trips, not foraging efficiency (which Tenczar didn’t measure).
If efficiency comes with experience – as suggested – it may be that additional time would also be needed to turn these extra flights into foraging trips that significantly benefitted the colony.
All of which means that stressors that adversely affect ageing bees, or that shorten the lifespan of foragers, may have a marked impact on colony pollen and nectar collection.
And there are lots of these sorts of stressors … pesticides, pollution, poor nutrition and pathogens – an alliterative gamut of threats to these important, elderly but highly effective bees. For example, increasing cumulative exposure to sub-lethal levels of pesticides may be deleterious to older bees.
Deformed wing virus
Unsurprisingly, being a virologist, it is the pathogens that interest me. In particular, deformed wing virus (DWV).

DWV symptoms
DWV is probably responsible for the majority of overwintering colony losses because it reduces the longevity of the (nominally long-lived) winter bees. I’ve discussed this at length elsewhere but the important bits are as follows:
- winter bees should live for months, not weeks, maintaining the colony through until springtime
- if there are lots of Varroa present during the early autumn (when winter bees are being reared) the developing winter bees will have high levels of DWV
- some bees will die before emergence, but those that don’t will instead die in weeks, not months
- consequently the winter cluster shrinks rapidly in size, becomes unable to thermoregulate and is separated from its stores
- this doesn’t end well … the colony either dies, or struggles through to the spring and is too weak to expand
But what happens to foragers with high levels of DWV in the summer?
Studies from my lab {{10}} have shown that pupae injected with DWV – essentially recapitulating what Varroa does when it feeds on a developing pupa – have three potential fates; they either die during development (~15%; see B in the figure below), emerge with developmental abnormalities (~65%; the deformed wing bit which ’does what it says on the tin’) or emerge and appear ‘normal’ (~20%).
Unanswered questions
Interestingly, the small proportion of bees that appear ‘normal’ have indistinguishable levels of DWV to those that have deformed wings (panel A below).
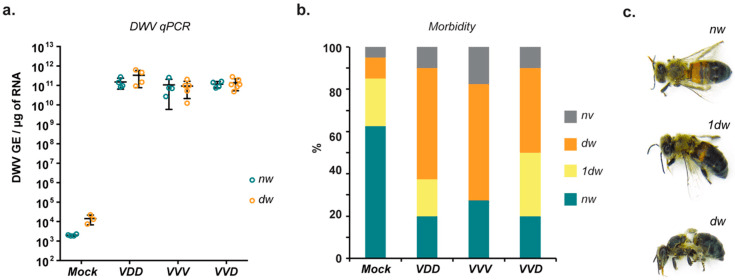
The fate of bees injected with DWV
Do these bees with high-DWV levels live long enough to become foragers?
We don’t know.
If they do become foragers – which , frankly, I doubt – do they learn how to forage well?
Again, we don’t know, though we do know from other studies that high levels of DWV leads to some cognitive impairment, so navigation at least may well be suspect {{11}}.
Finally, if they do become foragers, do they live a long and healthy life, or do they die prematurely?
Unfortunately … we don’t know this either.
Hive inspections
That’s a lot of ‘ifs’ there … and just as many unanswered questions.
Let’s assume that bees with high levels of DWV can mature to become foragers, but that they exhibit reduced longevity. If that is the case then the elite forager population would be reduced, so jeopardising provisioning the colony with nectar and pollen.
I’m sceptical that bees with such high DWV levels can survive long enough to mature into elite foragers. Nevertheless, I’d prefer to test this experimentally in the comfort of my lab {{12}}, rather than in my honey-production or queen-rearing colonies.
Therefore, during hive inspections I look carefully for the signs of overt DWV disease or varroosis – bees with deformed wings, uncapped developing brood or phoretic mites. I also periodically measure mite drop. If I see see problems (and with correct timing and appropriate treatment in autumn and winter you shouldn’t {{13}} ) I intervene.
Midseason mite management may save the elite foragers … and help prevent the loss of the colony overwinter.
A gradation of DWV levels
However, there’s a related – more subtle – thing to consider.
Studies from our group (and others) have shown that injection of tiny amounts of DWV results in a very rapid replication of DWV to stratospherically high levels. As shown above, these kill or maim ~80% of exposed bees.
But there’s a less well understood feature of colonies with high Varroa levels. During the course of the season the levels of DWV in bees not exposed to Varroa during development rise.
In March or April, DWV levels may be ~103/bee {{14}}. This is about the lowest level we ever see in bees, and is equivalent to the levels of DWV present in colonies from Varroa-free regions like Colonsay.
However, by mid- or late-summer the levels are 100-1000 times higher i.e. 105-106/bee {{15}}. This is still 10,000 times lower than the levels DWV reaches in Varroa-parasitised pupae {{16}}.
As an aside, we don’t formally understand how the DWV levels increase during the season. I suspect it’s through trophallaxis though we have also published some evidence of larval susceptibility to DWV during feeding.
Whatever the mechanism, bees carrying one million copies of DWV look completely normal and, as far as we can tell, behave completely normally.
‘As far as we can tell’, as no one has really done the right experiments …
I think it would be very interesting to carefully investigate the longevity and ability to achieve elite forager status for early season (very low DWV levels) and midsummer (intermediate DWV levels) bees.
Perhaps these intermediate levels of DWV are damaging after all?
Note
The Klein et al., paper has been very poorly proofread {{17}} and contains several errors, some of which potentially change the meaning of the text.
{{1}}: Note to self … why haven’t I written about the biology of guard bees yet?
{{2}}: At least it’s termed that by scientists … anyone sensible just says they do ‘different jobs at different times’. For convenience I’m going to skip over the plasticity in this process – the fact that they can be reversed, or that particular stages can be stalled for weeks or even months.
{{3}}: Be warned, they have a woeful implementation of rapidly refreshing embedded adverts which makes the page jump about infuriatingly … here’s a PDF copy of the page ‘frozen in time’ without the adverts.
{{4}}: Tenczar et al., (2014) Automated monitoring reveals extreme interindividual variation and plasticity in honeybee foraging activity levels. Animal Behaviour 95:41-48 https://doi.org/10.1016/j.anbehav.2014.06.006.
{{5}}: Who remembers this from the 1970’s? … “Steve Austin, astronaut. A man barely alive. Gentlemen, we can rebuild him. We have the technology. We have the capability to build the world’s first bionic man. Steve Austin will be that man. Better than he was before. Better, stronger, faster.” The Six Million Dollar Man … OK, you had to be there.
{{6}}: It might work perfectly well for inventory management in Marks and Spencer’s but detecting a 1 mm2 tag on a bee was a different matter.
{{7}}: My use of the term ‘lazy’ is sloppy anthropomorphism … maybe these should be termed inferior foragers?
{{8}}: Klein, S., et al. (2019) Honey bees increase their foraging performance and frequency of pollen trips through experience. Sci Rep 9: 6778. https://doi.org/10.1038/s41598-019-42677-x.
{{9}}: In fact, using a similar analysis method as Tenczar et al., – which defined elite bees in terms of individual activity compared to total colony activity – Klein calculate that just 12% of the bees contributed 50% of the foraging trips.
{{10}}: Gusachenko, et al., (2020) Reverse Genetic Analysis of Deformed Wing Virus Transmission, Replication, and Tropism. Viruses 12:532. https://doi.org/10.3390/v12050532
{{11}}: There are studies showing that foraging is impaired when adult bees are injected with DWV, though the virus levels were highly variable and, importantly, the virus wasn’t present during key developmental stages.
{{12}}: We’re not doing this as we’re too busy doing other things. If anyone wants to offer …
{{13}}: And I generally don’t, and haven’t this year.
{{14}}: 1000 copies, for those unfamiliar with scientific notation; we actually quantify them in a different way but I’ve simplified the units.
{{15}}: 100,000 to one million copies.
{{16}}: 10 billion copies (or higher!).
{{17}}: Yes, even worse than this post.
Join the discussion ...