You are what you eat

Honey bees are eusocial insects. The ’eu’ before social is derived from the Greek for ‘good’; these insects – not just honey bees – exhibit the highest level of social organisation, involving the division of labour (nurses, scouts, guards, foragers and in reproductive and non-reproductive sub-groups), cooperative brood rearing and overlapping generations.
Eusociality is a response to evolutionary pressure. Colonies are more successful than individuals.
Sociality involves cooperation. This requires recognition of kin and/or other group members. It also involves defending the shared resources that the group have worked together to collect (stores) or rear (brood).
Without defence, all that hard work would be lost to other competing groups who would try and steal the resources.
Social groups have therefore developed ways to distinguish themselves from other social groups of the same species. Although some social insects use visual clues {{1}} to distinguish group members (Baracchi et al., 2015), the majority use odours, in particular cuticular hydrocarbons (CHC’s).
Insects probably first evolved the ability to produce these hydrophobic CHC’s as waterproofing agents. However, they have subsequently been co-opted into use as pheromones for communication and, in honey bees, as a means of distinguishing nestmates from robbers (or, let’s be generous here, non-nestmates 😉 ).
Diversity
There is a huge volume of (sometimes contradictory) studies on CHC-mediated nestmate recognition. I’m not qualified to provide a comprehensive review. Instead, I’m going to focus on a couple of recent studies that appear to have resolved some confusing aspects of CHC’s and nestmate recognition.
Whilst this may sound rather dry and academic ((It isn’t, honestly.)), I will try and frame it in the context of practical aspects of honey bees of relevance to beekeepers, in particular the diversity of bees in the hive and the activities of robbing and drifting.
I’ll discuss these things first before I bore you to tears with the science 😉 .
Diversity in the colony – by which I mean genetic diversity – is critical for survival. The now classic studies by Mattila and Seeley (2007) demonstrated that queens that mated with more drones headed colonies that were ‘fitter’. Colony fitness was manifest by an increased behavioural repertoire of workers, a reduced production of diploid drones (which would otherwise be produced due to the single-locus sex determination system) and an increased resistance to a wide range of parasites and pathogens.
Genetic diversity is not only a good thing, but is essential for colonies to flourish.
I’ve discussed diversity previously together with the related concept of polyandry … (for once) I won’t repeat myself. And, while you’re there, you might enjoy Who’s the Daddy, which is related to hyperpolyandry, and demonstrates that bees preferentially choose the rarest patrilines when selecting larvae for emergency queen rearing {{2}} which may act as a mechanism to maintain diversity.
Drifting
Beekeepers understand the wonderful ‘homing’ abilities of their bees. They can fly off and forage miles away and return – apparently unerringly – to the hive.

Laden foragers returning …
It therefore comes as quite a surprise when beekeepers are told that a very significant proportion of the bees in a hive may not have been reared in the that hive {{3}}.
And by ’very significant proportion’ I mean up to 40%.
That’s right, 4 out of every 10 workers may well have been reared in another hive. In a colony containing 30,000 workers, 12,000 of them may have started life in a different colony.
We know this because Pfeiffer and Crailsham (1998) conducted studies using large numbers of colour marked bees and showed that, over time, they increasingly appeared in adjacent hives. The overall proportion that ‘drifted’ was dependent upon the position of the hive and the time of season.
In fact, canny beekeepers have long known about drifting.

How to encourage drifting …
Hives at the downwind end of row are supposed to accumulate more bees, and get stronger over time. It’s worth noting that this ‘fact’ is contradicted by the Pfeiffer and Crailsham studies who showed that it was hives in the middle of a row that received the most bees by drifting.
Whatever … back to drifting.
Drifting is thought to be predominantly, though not exclusively, due to young flying bees going astray on their orientation flights. The very fact that up to 40% of the adult bee population may have drifted in from elsewhere suggests there may be little control over it, or even mechanisms to stop it.
Robbing
Finally, the other relevant behaviour I want to briefly discuss is robbing. This is where the foragers from one colony {{4}}, rather than collecting nectar from clover or the flowering lime, instead rob the stores from another colony.
The guard bees in the colony being robbed are overwhelmed by the large numbers of robbers. This can be very distressing for the beekeeper and very destructive for the robbed colony.
Unlike drifting, which is essentially a short-range phenomenon, robbing can occur over much greater distances. I’ve previously tracked robbing bees back across three fields in Warwickshire – perhaps 1 km – and it probably occurs over even longer ranges.
Robbing is a problem for the colony being robbed, and may be a problem for the colony doing the robbing.
One reason a colony gets robbed is that it is weakened by parasites or disease. For example, a colony with unmanaged and high levels of Varroa will have very high deformed wing virus levels, and a large proportion of workers will emerge with wing deformities (or die before emergence). When robbed, the phoretic Varroa mites hitch a lift back to the robbing colony. Crime doesn’t pay.
Contradictions and guard bees
Some of you may have noticed an apparent contradiction in the paragraphs above.
Drifting bees appear to be able to relatively easy enter a different hive, whereas robbing only occurs when the guard bees are ‘overwhelmed’ by the robbers.
Of course, since it’s bees and beekeeping, it’s not quite that simple 🙂 .
There is ‘silent’ robbing where the guard bees appear not to defend the colony, and many beekeepers are aware that nectar-laden foragers {{5}} – typical of a really good nectar flow – may easily enter hives other than their own.
I think I can explain drifting bees on orientation flights (and will in due course), but let us first consider guard bee activity … this is supposed to prevent robbing.
Robbing is worse when there is a dearth of nectar.
Under these conditions foragers cannot find the nectar they want from flowers or trees, so instead try and obtain it from other colonies. If successful, the robber subsequently uses the waggle dance to recruit more foragers which, in turn, attempt to rob the targeted colony.
It can then quickly get out of control.
Guard activity is inversely related to nectar availability
Downs and Ratnieks (2000) studied honey bee guard activity during a period in which environmental nectar availability changed. The latter was cleverly monitored by recording the minimum syrup concentration that foragers would collect each day.
During a dearth the bees would happily visit feeders containing weak syrup, but during a strong nectar flow they would ignore weak syrup and only collect very strong sugar solutions {{6}}.
Hive weights increased as the bees showed a preference for stronger and stronger syrup, reflecting nectar being stored.
At the same time, the scientists were recording guard bee numbers and fights at the hive entrance. As the hive weight increased these both reduced; during strong nectar flows there are fewer guard bees and those present don’t defend the hive as strongly as during a nectar dearth.
But the most telling experiment was challenging hives with introduced bees from the same or different hives (nestmates or non-nestmates as they’re described in the paper).
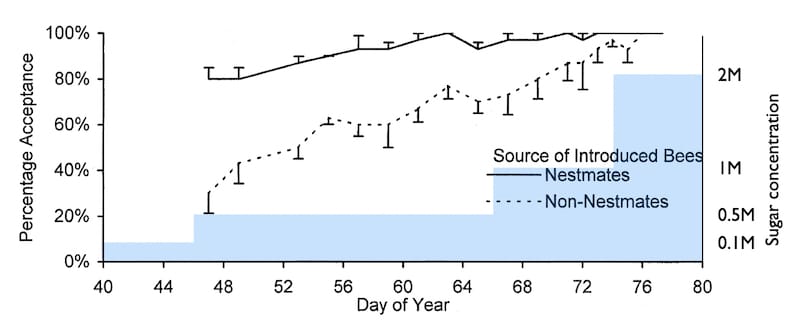
As nectar availability increases (indicated by blue shading) guard bee activity decreases
During a nectar dearth (before day 50) the guards accepted 80% of introduced nestmates but only 25% of non-nestmates. However, as nectar availability increased guards became more permissive to both nestmates and non-nestmates.
Eventually an ‘accept all’ threshold was passed (after day 74) and the guards let everyone in.
So, guarding activity is inversely related to environmental nectar availability. There’s some other interesting stuff in this paper, including things on the guards to fights ratio, but that’s for another time.
I’ll return to these acceptance percentages of nestmates and non-nestmates later, but first let’s consider how the guards distinguish friend from foe.
CHC’s and genetic relatedness
CHC’s are long-chain hydrocarbons (predominantly alkanes, alkenes, and branched alkanes) made in special cells called oenocytes. After synthesis they are transported to the outer cuticle of the insect where they form a water-repellent layer that helps prevent desiccation {{7}}.
CHC’s are also pheromones and the ratios of the various hydrocarbons in the mix differ between colonies, providing a unique ‘fingerprint’ that allows guard bees to discriminate between nestmates and non-nestmates.
The majority of the bees in a colony are genetically related as they share the same mother (the queen). Although there may be 15-30 (or 70) drones that have also contributed, the shared maternal genes – the workers are all half-sisters – could provide the unique genetic identity that accounts for these differences in CHC’s.
But what about all those genetically distinct bees that drifted into the colony from elsewhere?
I’m pleased you asked … you’re obviously paying attention 😉 .
Environment and ‘Gestalt’ theory
The presence of large numbers (up to 40%) of non-nestmates within a colony is compelling evidence that genetic differences are not the only, or even the primary, determinant of the variation in CHC profiles between colonies. If they were, drifting bees would always smell different and would be rejected at the colony entrance 75% of the time (during a nectar dearth – see above).
They might get in once (25%), but repeated flights from the hive would mean they would quickly be excluded; twice (~6%), three time (~1.5%) etc. If that were the case non-nestmates could never reach 40% of the hive population.
There has to be another explanation.
Perhaps there are ‘hive smells’ that are unique and are acquired by the bees within a colony. If this were the case non-nestmates would, after drifting into the colony, acquire the colony-specific CHC profile and so subsequently be accepted by the guard bees.
This is essentially what the ‘Gestalt’ theory says …
Colony-specific odours are transferred and homogenised between individuals within the colony, either directly (for example by contact or feeding) or indirectly.
Michael Breed published several papers in the 1980’s that indicated a role for wax comb as mediating transfer of these odours between individuals in the colony.
However, whilst the evidence for this Gestalt theory is reasonably compelling for some ant species, it doesn’t appear to fully explain what happens with honey bees (or, for that matter, many other social insects).
But two recent studies from scientists at the Washington University in Saint Louis have made some important and interesting contributions that explain many of the features of the recognition/defence processes observed in honey bees.
Worker maturity and CHC profiles
It had been known for a long time that newly emerged bees have lower levels of CHC’s. Vernier et al., (2019) investigated this further and showed that the range of CHC’s expressed changes from emergence and only stabilises once a worker leaves the hive and becomes a forager.
As a reminder, workers spend about 3 weeks as hive bees, cleaning cells, feeding larvae, supporting the queen, honey processing etc. before taking on roles that involve leaving the hive such as undertaker bees, scouts and – finally – foragers. Foragers are the oldest bees in the hive and normally die ‘in the field’.
The CHC profile undergoes a series of modifications as the worker transitions through these usually age-related behavioural tasks. Overall CHC levels reach a stable maximum two weeks after emergence, but the levels and ratios of individual hydrocarbons within the total population change as the bee matures.
It is only the foragers that exhibit the full colony-specific CHC profile.
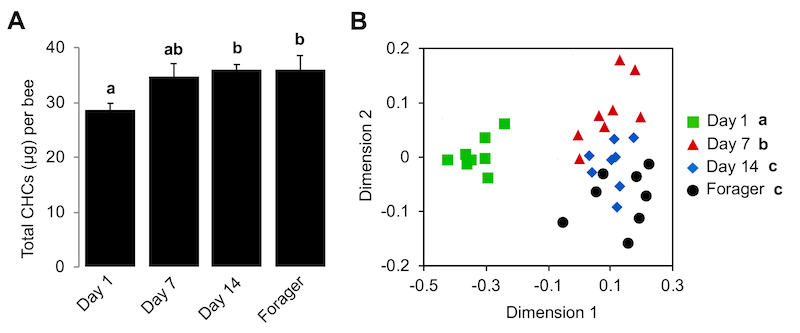
CHC levels (A) and profiles (B) in bees from a single hive
What’s more, these CHC profile changes are task-related, but not age-related. The scientists established small colonies containing genetically-related bees of the same age. Under these circumstances, some bees rapidly mature into foragers, whereas others remain as nurses … despite being the same age.
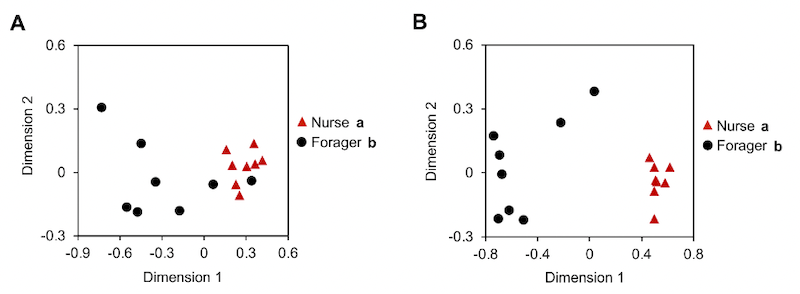
Bees of one week (A) or three weeks (B) of age exhibit role-specific CHC profiles
Finally (to truncate an excellent data-rich paper in the interests of completing this post in less than 22,000 words) the authors demonstrated that the final maturation of the CHC profile required that the worker leaves the hive. This suggests that there is an environmental contribution of some sort. If older foragers were ‘forced’ to revert to nurse bee roles they retain their mature CHC colony-specific, forager-like profile.
Statistics aside …
The CHC profile of an individual bee is defined by the levels of 19 different hydrocarbons measured by gas chromatography. Since each hydrocarbon may differ in abundance it is very difficult to ‘see’ patterns, so making comparisons tricky. For this reason a statistical method termed nonmetric multidimensional scaling was used which allows the visualisation of clusters of similar data points – as seen in three of the four graphs above. Just look for the patterns … if all the similar symbols are clustered together (below, left) this means that the CHC profiles are distinctly different. If the symbols are all jumbled up together (below, right) then it’s not possible to discriminate between the CHC profiles.
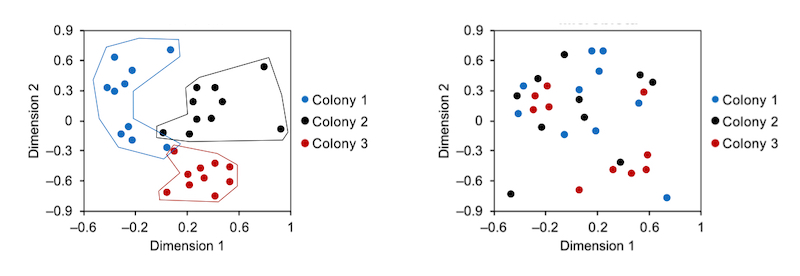
Three reasonably distinct clusters (left) and unclustered data (right)
Exactly the same analysis method is applied to the bacterial population in the honey bee gut … but I’m getting ahead of myself.
With apologies for the above to any statisticians reading this 😉 .
Drifting bees and CHC profile maturation
Although it’s not discussed in this Vernier et al., paper, I wonder whether the maturation of the CHC profile they describe – only achieving a colony-specific identity in foragers – explains why bees on orientation flights are readily accepted when they drift to the wrong colony?
My expectation would be that these bees, on their first flights from the hive, have yet to achieve the stable, mature and colony-specific CHC profile. They would therefore look smell much more similar to younger-than-forager bees from any colony, including the one that the guards were there to protect.
Presumably guards have evolved methods to not reject nestmates on their first orientation flights, it would be self-defeating if they didn’t.
If I’m right, non-nestmate bees on orientation flights would smell ‘OK’ and be let in, but older foragers – with the mature and different colony-specific CHC profiles – would be rejected.
This needs formally testing … I’m just thinking out loud.
I would look at rejection rates and CHC profiles of bees on orientation flights and compare them to age-matched nurse bees or foragers from the same cohort. I would expect it would only be the bees actively involved in foraging that would be rejected.
But what determines the differences in the CHC profiles if it isn’t genetic variation between bees? And it isn’t, or bees acquired by drifting would never be able to make successive flights as explained above.
You are what you eat
In another paper, Vernier et al., (2020) have at least a partial explanation for the colony-specific CHC profile by which nestmates are distinguished from non-nestmates.
It is due to the range and – more importantly – the numbers of each species and strain of bacteria in the honey bee gut. Together, this microbial population is called the microbiome, specifically the gut microbiome.
The authors demonstrate that foragers from different colonies have different microbiomes (A, below) and distinctively different CHC profiles (B). However, tellingly, by transferring newly emerged bees between colonies, they demonstrated that the resulting microbiome largely (though not exclusively) reflected the colony the bee was present in post-emergence (E).
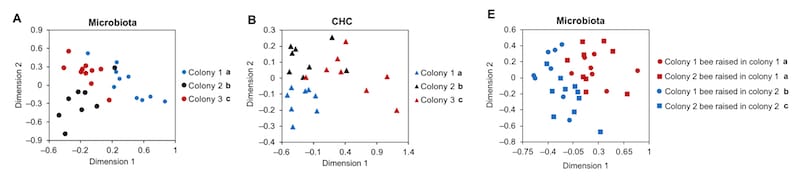
Microbiome (A) and CHC profile (B) differ between colonies, but CHC profile depends upon forager environment (E)
Antibiotics can be used to kill the bacteria in the bee gut. Antibiotic-treated bees had fundamentally altered microbiomes and, more importantly, CHC profiles.
In addition, because newly emerged bees acquire their microbiome during feeding by older bees in the hive, they showed that antibiotic treatment of older bees altered the resulting microbiome of the newly emerged bees they were co-housed with, irrespective of the hive the latter were actually born in.
Finally, Vernier et al., (2020) manipulated the microbiome by inoculating bees with strains of Gilliamella apicola, a naturally present symbiont in the bee gut involved in sugar metabolism. Inoculated bees had altered microbiomes {{8}} and altered CHC profiles.
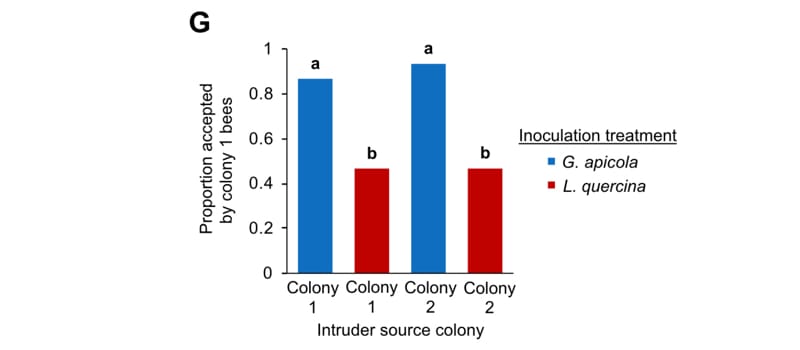
Guard acceptance after microbiome manipulation (L. quercina is an environmental negative control inoculum)
Even more compellingly (as what we’re really interested in is guard bee recognition and rejection) bees inoculated with Gilliamella apicola were more likely to be accepted by guard bees harbouring the same strain of bacteria, irrespective of the genetic origin of the foragers being tested.
So, in summary … just in case you dozed off
Guard bees protect the hive, particularly in times of nectar dearth, and discriminate between nestmates and non-nestmates by the complex profile of CHC’s they are coated with. You can see guard bees using antennation to ‘smell’ returning foragers if you look closely at the hive entrance.
Mature foragers are usually rejected if they are from a different colony, at least during a dearth. If there’s a strong flow on, it’s a free-for-all.
Lots of bees move between colonies and most of this happens during the orientation flights taken when the bee first leaves the hive (about 3 weeks post-emergence).
The CHC profile of a bee changes and matures as the bee takes on different roles after emergence. It is only mature foragers that have a fully mature CHC profile. I suggest above that bees on orientation flights might have yet to acquire this mature forager-like CHC profile.
The gut microbiome, acquired in the hive by transfer from older bees, is the primary determinant of the CHC profile finally exhibited by the forager. Clearly this needs to change and mature somehow during the time the bee spends in the hive after emergence and before becoming a mature forager.
How this occurs is unclear; are they fed different things at different times (for example, by different cohorts of older bees), or does the microbiome change in response to their diet, or perhaps even the ‘job’ they are doing? {{9}}
Unanswered questions
This is lovely science and, although it has little to do with beekeeping per se, it helps explain some of the characteristics our bees exhibit e.g. why orientating bees drift so freely and are not rejected.
However, like all science, it creates a lot more questions that have yet to be answered.
- How does the microbiome influence the CHC profile? The CHC’s are produced by specialised cells called oenocytes and these are not directly exposed to the microbiome. We know that the changes in CHC profile are due to different levels of gene expression in the oenocytes, so it seems likely that metabolites produced (or used up) by the microbiome influence this gene expression.
- How did this process evolve? It turns out other social Hymenoptera also share certain features of this microbiome/nestmate recognition process, suggesting a common – and therefore probably ancient – evolutionary origin. It is a mutually beneficial interaction; the host (bee) acquires a mechanism for ‘social recognition’ of nestmates, but the bacteria also benefit by protection and preventing invasion by other hosts bearing different gut bacteria. This is more theoretical than practical, but interesting nevertheless {{10}}.
But what about?
In reading some of the background literature, I’ve also got some slightly more practical bee-related unanswered questions:
- Are the 80%/25% acceptance ratios of nestmates/non-nestmates correct? Rejecting 20% of foraging nestmates during a dearth seems verging on suicidal. Surely this limits nectar collection? Likewise, allowing 1 in 4 non-nestmates in seems a bit risky. Perhaps these figures are influenced by the experimental design. With barcoding/RFID methods this might be worth revisiting.
- Remember the elite foragers? These are the 20% of foragers that account for 50% of the foraging activity of the colony. Are the rejection/acceptance rates of elite foragers different from their more work-shy half-sisters? Elite foragers not only make more trips, but also collect more pollen, nectar or water on each trip.
- Oxytetracycline (OTC) may be used to treat European foulbrood in the UK {{11}}. The antibiotic kills the causative agent, Melissococcus plutonius. However, it will also slaughter a proportion of the gut microbiome. What happens to guard bee activity and nestmate recognition in a tetracycline-treated hive? There are already studies of the impact of OTC on the microbiome … and on the microbiome transferred to newly-emerged bees (Jia et al., 2022). I suspect the CHC profile and nestmate recognition would inevitably change as well … and possibly not in a good way.
Notes
Yikes! Like Topsy, that ‘growed’ rather more than I intended it to. Congratulations to those who made it to the end. There are only so many preparing for the season ahead-type posts that are possible without repetition, so I thought some bee-related science was in order. What’s more, with the weather about to turn a whole lot worse, the start of the season is still quite a way off. Any of you who are already feeding thin syrup – and there will be – might want to check whether it’s frozen solid in the rapid feeder 😉 .
Finally, to the correspondent who wrote and asked whether he could split his colony in early March … don’t, just don’t.
References
Baracchi, D., Petrocelli, I., Chittka, L., Ricciardi, G., and Turillazzi, S. (2015) Speed and accuracy in nest-mate recognition: a hover wasp prioritizes face recognition over colony odour cues to minimize intrusion by outsiders. Proceedings of the Royal Society B: Biological Sciences 282: 20142750 https://royalsocietypublishing.org/doi/10.1098/rspb.2014.2750.
Downs, S.G., and Ratnieks, F.L.W. (2000) Adaptive shifts in honey bee (Apis mellifera L.) guarding behavior support predictions of the acceptance threshold model. Behavioral Ecology 11: 326–333 https://doi.org/10.1093/beheco/11.3.326.
Jia, S., Wu, Y., Chen, G., Wang, S., Hu, F., and Zheng, H. (2021) The Pass-on Effect of Tetracycline-Induced Honey Bee (Apis mellifera) Gut Community Dysbiosis. Front Microbiol 12: 781746.
Mattila, H.R., and Seeley, T.D. (2007) Genetic Diversity in Honey Bee Colonies Enhances Productivity and Fitness. Science 317: 362–364 https://www.science.org/doi/10.1126/science.1143046.
Pfeiffer, K.J., and Crailsheim, K. (1998) Drifting of honeybees. Insectes soc 45: 151–167 https://doi.org/10.1007/s000400050076.
Vernier, C.L., Chin, I.M., Adu-Oppong, B., Krupp, J.J., Levine, J., Dantas, G., and Ben-Shahar, Y. (2020) The gut microbiome defines social group membership in honey bee colonies. Science Advances 6: eabd3431 https://www.science.org/doi/10.1126/sciadv.abd3431.
Vernier, C.L., Krupp, J.J., Marcus, K., Hefetz, A., Levine, J.D., and Ben-Shahar, Y. (2019) The cuticular hydrocarbon profiles of honey bee workers develop via a socially-modulated innate process. eLife 8: e41855 https://doi.org/10.7554/eLife.41855.
{{1}}: Literally ‘face recognition’ in hover wasps Liostenogaster sp.
{{2}}: It’s a particularly intriguing story.
{{3}}: I used to test this with a ‘show of hands’ during in-person talks … it is officially a ‘little known fact’.
{{4}}: Or, even worse, several colonies.
{{5}}: i.e. experienced bees not on orientation flights.
{{6}}: Foragers determine the sugar content of nectar and conduct a cost/benefit analysis of collecting it or flying further to a better source.
{{7}}: They stop water getting out.
{{8}}: Big surprise, not!
{{9}}: That last one would be very cool.
{{10}}: At least, it is to me!
{{11}}: Though shook swarming the colony is now the preferred approach I think. In other countries different regulations apply.
Join the discussion ...