Socially distanced bees
A real skill when writing scientific papers {{1}} is to give them a suitable title.
Choosing the title involves a combination of art and science.
It must look appealing … you want the viewer to become a reader.
Since it is always indexed by search engines you must make sure it includes suitable keywords or phrases.
It needs to be informative. At least sufficiently so that the ‘take home message’ is clear. Even if the viewer does not become a reader they should still remember the title and so know the gist of what the article concludes.
The art of good title writing goes beyond this though. To increase the appeal, if it includes humour, some sort of half-hidden pun or some clever word play, then all the better.
And there are some great examples out there:
- You probably think this paper’s about you: narcissists’ perceptions of their personality and reputation by Erika Carlson et al. (2011) in Journal of personality and social psychology 101:185-201. doi:10.1037/a0023781
- Fifty ways to love your lever: Myosin Motors by Steven Block (1996) in Cell 87:151-157 https://doi.org/10.1016/S0092-8674(00)81332-X
There’s another variant of the latter and a host of additional variously funny or insensitive titles in this post on Slate. This also includes mention of the contrived efforts some scientists make to include Bob Dylan song titles in their publications (see Freewheelin’ scientists: citing Bob Dylan in the biomedical literature in the BMJ) as part of a long-running bet with colleagues.
Making it topical
Failing humour – and you could argue that some of the examples above {{2}} or linked are failing humour – a good way to get a paper some attention is to use a title that overtly hints at topicality.
In this regard, two papers caught my eye {{3}} this week:
- No time to die: An in-depth analysis of James Bond’s exposure to infectious agents by Wouter Graumans et al., (2021) in Travel Medicine and Infectious Disease 44:102175 https://doi.org/10.1016/j.tmaid.2021.102175
- Honey bees increase social distancing when facing the ectoparasite Varroa destructor by Michelina Pusceddu et al., (2021) in Science Advances 7:eabj1398 https://www.science.org/doi/10.1126/sciadv.abj1398
The first of these is topical because travel restrictions to limit infectious disease transmission is a near-daily news item. However, it goes further than that in also including the Blofeld-like quote. The paper also has an entertaining abstract which finishes with the words We only live once, and sub-sections entitled The man with the golden gut: food safety and infections and The fly who loved me: arthropod-borne diseases.
However, I’m not going to discuss the analysis of Bond’s hand-washing, potential Toxoplasmosis or the disturbingly high mortality rate of his sexual partners.
You’ve seen the film(s), now read the book paper 😉
Instead I’ll briefly focus on the second paper which managed to sneak ‘social distancing’ into the title, thereby ensuring it was picked up by almost every newspaper in the UK.
Socially distanced bees
‘Briefly’ because it’s a long paper and because rather too many of the figures are uninspiring bar charts like this one:
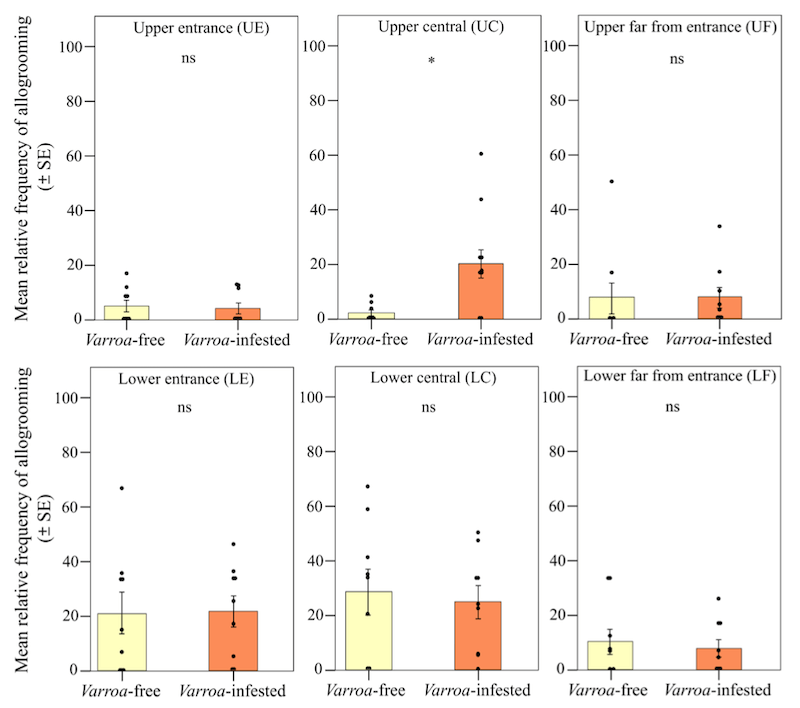
Spatial shift in allogrooming behaviour
… which, if you read the legend shows that there is almost no significant (ns) difference in allogrooming behaviour (which I’ll come to shortly) between Varroa-infested and -uninfested bees.
However, some of the graphs do have bars of different heights (and that are statistically significantly different) and there’s an interesting contradiction between studies conducted on full colonies and individual cohorts of bees.
So, rather than work through the entire paper I’m going to just focus on a few points and then discuss a couple of things that I found interesting.
Hypothesis driven science
Social insects, like ants and bees, are particularly at risk from pathogens and parasites. Their large populations, high density and ample food reserves means they have had to evolve both individual and social immunity.
The former prevents or mitigates infection of the individual, the latter reduces the chances that the colony will get infested (or restricts the impact of any infestation or infection to help ensure the survival of the colony).
The authors hypothesised that the presence of Varroa might induce some of these social immune responses. For example, bees might increase grooming activity in areas of the hive where Varroa were most frequent, or they might decrease antennation or trophallaxis with infested nest-mates, all to reduce the chance of mite transmission.
They focused on two particular aspects of social immunity and colony organisation, and made two predictions (hypotheses) for each:
- Space usage.
- Spatial shift of waggle dances to the periphery of the brood nest in infested colonies when compared with uninfested colonies.
- Spatial shift of grooming activity to the core of the colony in infested colonies when compared with uninfested colonies.
- Social behaviour.
- Infested bees would be expected to show changes in social behaviour including an increase in allogrooming, and decreases in antennation and trophallaxis.
- Changes in the structure of the social network in the infested hive, with decreases in connectivity and centrality.
Using colonies with high and low (almost negligible – I’ll return to this later) mite levels they then conducted observational science – they watched waggle dances, allogrooming etc. – to see if their predictions were correct.
Compartmentalisation of the colony
When we open a hive all we often see is a mass of bees covering every frame.

Lots of bees
Beekeepers are often too busy trying to find the queen, or judge whether there are eggs or sufficient stores present, to appreciate that the bees are organised into two main ‘compartments’ within the colony:
- an outer one occupied by foragers (the older bees) located nearer the hive entrance.
- an inner one containing the young nurse bees and the queen, all of which are mainly arranged on brood.
The authors reasoned that since foragers represent a potential entry route of Varroa into the hive, you might expect the waggle dancing foragers to move the ‘dance floor’ to the periphery of the colony.
Does this make sense to you? To me it only really makes sense if you assume that the forager picks up a mite from elsewhere, for example when robbing a mite-infested collapsing colony elsewhere and returning to the hive. The alternative is that that forager was already carrying a mite, though I suppose that’s still a mite being introduced (or, more correctly, reintroduced) to the colony
Whatever the reason – and this wasn’t really elaborated – the changes in space usage and social behaviour would be expected to increase the compartmentalisation of infested colonies, so reducing mite spread.
Remember, mites predominantly associate with nurse bees and need to spend several days ‘surfing’ around the colony on these bees before entering a cell to reproduce.
Experimental details
Two month before the experiments started observation hives and other colonies were treated with dribbled oxalic acid. The colonies destined to be “Varroa-free” were then treated once a week for two further weeks with trickled oxalic acid.
Six weeks later, at the start of the observations, Varroa levels were strikingly different. The infested colonies were about ~6.2% and the “Varroa-free” uninfested colonies ~0.1%.
6% means six mites for every 100 bees sampled.
The team recorded the location of waggle dances and allogrooming in observation hives. Independently, using individually marked populations of caged bees, they recorded allogrooming, antennation and trophallaxis.
And, just so we all know what these terms mean:
- allogrooming – is where one bee removes foreign particles and parasites from another bee
- antennation – is how bees identify nestmates in the hive, by touching with the antenna
- trophallaxis – is where one bee feeds another bee liquid food
Spatial shifts in waggle dancing and allogrooming
The colony is approximately spherical, sliced through by the vertically-hanging frames. The authors distinguished between the central frames and the lateral frames, and the position on the frames being closer or further away from the hive entrance {{4}}.
In uninfested colonies the waggle dance and allogrooming activity occurred on both central and lateral frames, and predominantly on the lower half of the frame.
In contrast, infested colonies showed a significant shift of waggle dancing activity to lateral frames, and to positions closer to the hive entrance on these lateral frames. The allogrooming activity also shifted, but in the opposite direction, becoming concentrated on a larger area of the central frame.
These spatial changes were statistically significant and they should have the effect of keeping the forager and nurse bee populations better separated, and of concentrating the grooming activity to the centre of the colony.

Spatial organisation of nurse bees (yellow) and foragers (red) in mite-infested and uninfested colonies
Did the latter occur because that’s where most of the mites are located … hanging around waiting for a suitably-aged late stage larva to snuggle up with?
Or, does allogrooming become concentrated in the core because the nurse bees – which are responsible for most allogrooming activity – have relocated from other areas within the colony?
Or both? … these are not mutually exclusive.
The diagram above is my half-assed rather poor attempt to demonstrate the changes in compartmentalisation within the colony. In the colony on the left there is much more mixing and overlap between the nurse and forager bees. On the right there is much less mixing, and therefore less opportunities for mite transmission.
Social behaviour
The studies on social behaviour were somewhat less definitive, or produced unexpected results. These studies were all done using caged bees from infested or uninfested colonies. Allogrooming, antennation and trophallaxis can all be divided into ‘giving’ and ‘receiving’ activity, all of which was recorded, as was whether the bee from the infested colony was activity carrying a mite.
The expectation was that these activities – all of which are likely to increase the opportunities for mite transmission – might all be reduced in bees from Varroa-infested colonies, with one or two caveats.
In fact, in the majority of cases there were no significant differences between the levels of allogrooming, antennation and trophallaxis.
The exceptions included Varroa-parasitised bees which were – perhaps understandably – more likely to be the recipients of grooming.
Infested colonies overall exhibited slightly increased antennation, with Varroa-carrying bees receiving significantly more attention from cage-mates and – in turn – performing less antennation.
Finally, although there was no overall difference between trophallaxis between bees from infested and uninfested colonies, bees actively parasitised by Varroa received more trophallaxis … an unexpected result considering the potential for mite spread.
The final hypothesis that was tested was whether the social network changed in infested colonies. This was based upon analysis of high resolution videos of caged bees, recording the interactions between and then calculating the connectivity and centrality of the network.
I’m deliberately being brief in my description of the methodology here, for two reasons; 1) it’s complicated and would take 500 words to describe more fully, and 2) there were no differences in the measured parameters of the social network in the infested bees when compared with the bees from the uninfested colonies.
Contradictions
Looking back at the predictions (see above) it seems clear that there were large scale changes in space usage within the colony … perhaps justifying the phrase ‘social distancing’ in the title.
However, when the authors looked at individual cohorts of bees they did not detect evidence of increased small scale separation – either within the social network they formed, or in terms of avoiding activities that would be expected to lead to mite transmission.
In fact, the caged bees showed increases in activities that were commensurate with ‘care giving’ … increased grooming and trophallaxis of Varroa-carrying individuals.
These appear to be contradictory observations.
How can the large scale spatial reorganisation occur without changes in the bee-to-bee interaction that occurs at a smaller scale?
The authors skirt around this a little, but don’t really tackle it head on.
Loose ends
I think a couple of things warrant further investigation.
The large scale spatial reorganisation was of activities (dancing and grooming) not of bees, though there was an unwritten assumption that the activities were observed to move because they were conducted by particular ages of bees (which did move).
That could be tested by high resolution video observations of a colony containing marked cohorts of nurse bees and foragers. The expectation would be that – like the red and yellow circles I’ve drawn above – you would expect to see a more distinct separation of the two groups.
With sufficient time, money and video recording you could also use this in place of the studies of small cohorts of caged bees. For example, using lots of bar coded bees. Perhaps these don’t perform in the same way outside the hive as inside it?
Oxalic acid treatment
The authors used oxalic acid to reduce mite levels in the “Varroa-free” hives.
Unusually – at least in my experience – they used three weekly treatments of trickled oxalic acid.
This seems to have been very effective in reducing mite levels – compare the 3 x treated (0.1% infestation) to the 1 x treated (>6% infestation) – five to eight weeks respectively after the treatment started.
I was surprised it was that effective in a colony that was activity rearing brood, where the majority of the mites would be hidden in capped cells.
However, there are numerous studies that show that trickled/dribbled oxalic acid damages open brood {{5}}. Therefore, in the studies conducted in this social distancing paper there’s a possibility that an entire generation of brood were missing due to the three successive treatments with trickled oxalic acid.
How this would have affected the results is unclear.
Although bees display temporal polyethism they also exhibit developmental plasticity and can change roles if and when needed. This doesn’t appear to have been considered and is certainly not discussed in the paper.
How is social distancing achieved?
But, let’s take their clever and topical title at face value and accept that bees do socially distance in response to mite infestation {{6}}.
What level of mite infestation is needed to initiate this activity?
What are the molecular (chemical) or behavioural signals that trigger this activity?
Can we, as beekeepers, exploit them to improve the efficacy of rational mite management?
All of which will involve wild speculation and precious few hard facts, so I’ll save it for another time 😉
{{1}}: And for that matter blog posts … something I have yet to master.
{{2}}: And definitely some on this site … for example Chewin’ the fat.
{{3}}: Actually, there was a third on crash landing bees and optic flow which I was hoping to weave together with a November 5th -related title of some sort “Crash, Bang blah blah” … but I ran out of time.
{{4}}: The frame were obviously arranged the ‘cold way’.
{{5}}: For example, see Terpin, B., Perkins, D., Richter, S. et al. A scientific note on the effect of oxalic acid on honey bee larvae. Apidologie, 50, 363–368 (2019).
{{6}}: Accepting also that it’s not really social distancing as we are all practising, but more like keeping separate the infected and uninfected … like flying the “Yellow Jack” or the introduction of strict quarantine in Ragusa in 1377 to restrict the Black Death.
Join the discussion ...